|
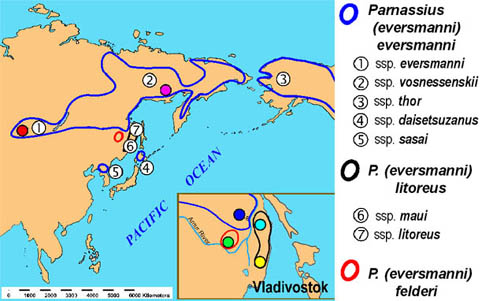
Introduction  Parnassius eversmanni, is a typical representative of alpine fauna in mountain ranges of Southern and Eastern Siberia. The species is also distributed on the vast territory of the Far East, it occurs in northern Mongolia, in northern Korea, in Japan (in Hokkaido), in northern USA (in Alaska), and north-western Canada (Korshounov, Gorbunov, 1995; Weiss, 1999). Extremely wide and at the same time disjunctive natural habitat of the species is the reason for its complex geographic variability (Fig. 1). It was the cause for describing numerous subspecies and forms in Parnassius eversmanni (Kurentsov, 1970) that were later grouped into two or three independent species (Weiss, 1991; Tuzov, 1993) or semispecies (Korshounov, Gorbunov, 1995; Glushchenko, Martynenko, 1998). | |
|
Fig. 1. Distribution of Parnassius eversmanni (s. lat.) and sources of the material used in our study (by Korshunov and Gorbunov, 1995 and Weiss, 1999). Filled circles indicate sampled localities. | |
|
Most of the known subspecies, although they often inhabit localities that are far apart, are not or are little differentiated. It is considered, that P. eversmanni inhabits most of Siberia and that there is a continuity from west to east with continual genetic exchanges between numerous local populations. Only the large subspecies from south-eastern Russia are more characterized and they still pose some taxonomic questions linked to the problems posed by Parnassius felderi (Fig. 2). This taxon, which essentially differs from the large forms of Parnassius eversmanni by having white wings instead of yellow, is reported as flying with this species in the district of Obluch'ye. According to other data, P. felderi and P. eversmanni do not cohabit but can be observed in localities that are close together. Finally, it is known that some specimens of both these taxa were collected in Kuldur. |
|
|
|
|
|
a |
b |
c |
|
Fig. 2. Morphological variability in Parnassius eversmanni : ssp. vosnessenskii (a), ssp. litoreus (b), ssp. felderi (c).
|
||



|
Thus, the problem is do we have two different species that are sympatric in the Khingan Mountains, or all these variable forms are simply the expression of a vast variability of Parnassius eversmanni in this region and therefore "felderi" is nothing more than a white morph of Parnasius eversmanni. The situation is complicated with two seasonal forms described in Parnassius eversmanni. In northern regions (Magadan, Yakutia) and Altai, there is normally a single biennial generation. But in south locations like southern Far East, early-summery and middle-summery generations are known. Butterflies of the early-summery generation are very similar to Parnassius eversmanni from Magadan both in wingspan and in intensity of dark pigmentation. Butterflies of the middle-summery generation differ in larger wingspan and weaker intensity of dark coloration. In spite of significant morphological differences, Yu.N. Glushchenko and A.B. Martynenko (1998) consider these two forms to be no more than just seasonal forms of the same species. The hypothesis is that some specimens in biennial population undergo an annual life cycle. As a result of such enormous variability, more than 40 infraspecific categories were described in Eversmann's Parnassian. But all the former attempts to revise intraspecific classification of this species were mainly based on the results of morphological studies and very rarely data on biology, ecology and phenology were taken into account. The aims of the present research were:
Materials and methods In present study, we used entomological material collected by Yu.N. Glushchenko and A.B. Martynenko in Primorye in the vicinities of Dal'negorsk, and in Khabarovsky region in the vicinities of Kuldur, Khingansk, Gorny and Vysokogorny. We also used the material from Yu.N. Glushchenko's private collection originating from Altai Mountains and from Magadan. At last, we used some specimens from A.I. Kurentsov's memorial collection, which is stored at the Institute of Biology and Soil Science FEB RAS. These specimens originated from Transbaikalia, Yakutia, Chukotka, Alaska, and Japan. Estimation of morphological peculiarities was performed by standard discriminant function analysis. Twenty-six characters were measured in 152 specimens. With the aim to estimate genetic variability in different parts of the natural habitat of Eversmann's Parnassian, a total of 77 specimens were tested. All the experimental procedures were carried out whether according to standard protocols or were optimized as was previously reported (Zakharov et al., 2000).
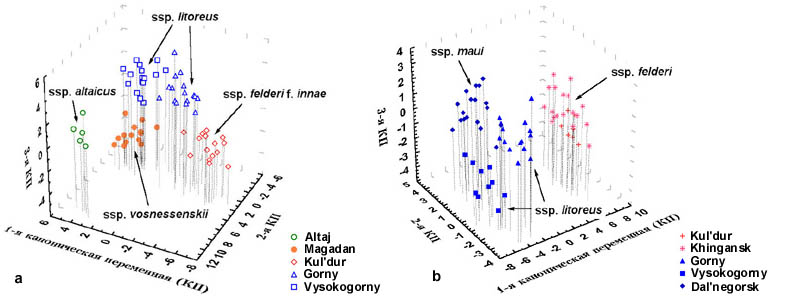
Results and discussion The discriminant model was calculated on the base of standard discriminant analysis (Fig. 3). These results are in principal agreement with data obtained by Glushchenko and Martynenko (1998). | |
|
Fig. 3. Distribution of early-summery (a) and middle-summery (b) specimens of Parnassius eversmanni in three-dimensional scale of principal canonical variables.
| |
|
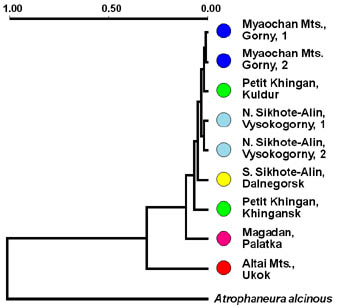
To estimate genetic diversity in Eight ten-base primers (OPA-04, OPA-07, OPA-08, OPA-10, OPA-14, OPA-18, OPB-20, and OPE-20) were used for RAPD-PCR. RAPD-patterns were aligned using RFLPscan computer program that produced binary matrix for further investigations. The results of this band-sharing analysis showed that distribution of genetic variability in Eversmann's Parnassian significantly differs in western and eastern parts of its natural habitat area. The specimens collected in Altai Mountains were the less polymorphic ones. All they had almost or absolutely identical RAPD-patterns, which were sharply different from those of Parnassius eversmanni from eastern parts of its natural habitat. The most complicated nature of genetic polymorphism was found in the populations in Primorye and in the south of Khabarovsky region. This is the territory, the greatest number of ecological subspecies and seasonal forms were described from. Due to high genetic variability, we were unable to identify any evident markers for individual populations. But two primers yielded characteristic patterns of amplification products for at least two samplings. Primer OPB-20 produced a 540 base pair major fragment, which frequently appeared in RAPD-patterns of the specimens from Kul'dur. The same fragment was rare among individuals from the vicinities of Gorny and Vysokogorny and was absent in RAPD-patterns of the specimens from Primorye, Magadan and Altai Mountains. These data correlate with the results of morphological survey made by Glushchenko and Martynenko. They revealed that specimens of Parnassius felderi with white wings are common in Maly Khingan but less frequently they also occur in Myoachan Mountain Range and northern Sikhote-Alin. Bright yellow wing coloration is characteristic for subspecies litoreus but some felderi specimens also have yellow tint. Another primer, OPA-08, produced a 440 base pair fragment that appeared to be major in the RAPD-patterns of all specimens from the vicinities of Dal'negorsk. The same fragment was found in RAPD-patterns of some other specimens of subspecies litoreus from the vicinities of Gorny and Vysokogorny. These locations are the most close ones to Dal'negorsk. A single primer OPE-20 was used to estimate genetic polymorphism among 14 individual museum specimens from Altai, Transbaikalia, Yakutia, Chukotka, Alaska, and Japan. As some specimens were collected as long ago as 1921, most DNA probes were heavily degraded, and we could not use these results in our general comparisons. But in this case, we have three specimens from Altai Mountains that have very similar patterns of amplification products. On the base of the binary matrix produced from general comparisons of RAPD-patterns, Nei and Li's genetic distances were calculated using RAPDistance freeware program. As it should be, specimens from Altai have the least average intrapopulation genetic distance - about 0.33. This value is almost the same with average intraspecific genetic distance based on RAPD data that was previously reported for Parnassius nomion, which is a well-diverged species. At the same time, this value is approximately two times lower than average intrapopulation distances calculated for eastern populations of Parnassius eversmanni. These results are in accordance with geographic isolation of the population from Altai Mountains. Cluster analysis of distance measures via UPGMA was carried out for individual specimens and populations. UPGMA phenogram plotted for individuals revealed no order in distribution of the specimens among populations. UPGMA phenogram based on average intrapopulation genetic distances does not show any evidences for genetic isolation among populations at the south of the Far East (Fig. 4). Moreover, it shows that middle-summery and early-summery generations sampled in two different localities are grouped according to the place of origin without any relation to flight periods. | |
Fig. 4. UPGMA dendrogram of genetic similarity between studied populations of Parnassius eversmanni.
|
|
|
With the aim to reveal more molecular markers to support RAPD-PCR data, we performed restriction analysis of 1600 base pair fragment of mitochondrial DNA gene coding cytochrome oxidase subunit I. DNA samples of 15 specimens from different populations were used as templates to amplify this fragment. Among 10 available endonucleases, three (AluI, MvaI, TaqI) were found to have restriction sites in the amplified fragment. As we can see, RFLP-PCR analysis revealed no variation among all tested specimens. Restriction analysis performed in ~2300 bp amplified fragments of mtDNA also revealed no variability among individuals and populations. Thus, we may conclude as follows:
References Glushchenko Yu. N., Martynenko A.B. (1998) Variability in parnassians of eversmanni-felderi species group in the southern Far East of Russia. A.I. Kurentsov's memorial meetings. Vol. VIII. Vladivostok. P. 41-56. [In Russian] Korshounov Yu.P., Gorbunov P.Yu. (1995) Butterflies of Asian part of Russia: Hand-book. Ekaterinburg. 202 p. [In Russian] Kurentsov A.I. (1970) The butterflies of the Far East USSR (Key). Leningrad, Nauka. 163 p. [In Russian] Tuzov V.K. (1993) The synonymic list of the butterflies from the ex-USSR. M.: Rosagroservice. 73 p. [In Russian] Weiss J.-C. (1999) The Parnassiinae of the world. Part 3. Canterbury: Hillside Books. 239 p. Zakharov E.V., Chelomina G.N., Zhuravlev Yu. N. (2000) Isolation and PCR with random and gene-specific primers of DNA of museum butterfly specimens of the family Papilionidae // Russian Journal of Genetics. Vol. 36. N 9. P. 1-9. |